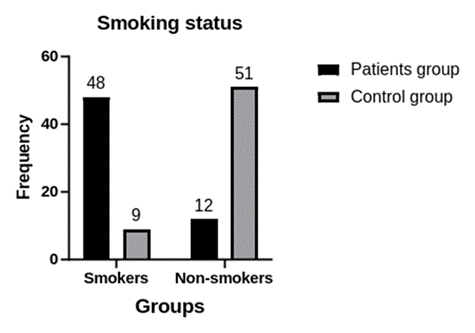
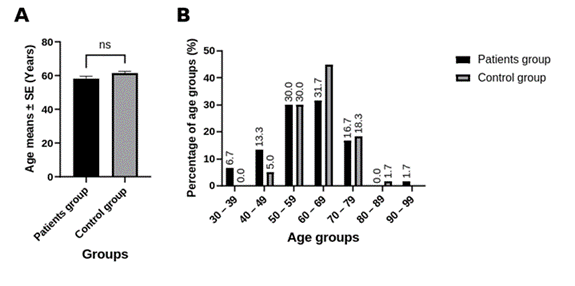
1. Introduction
Human metapneumovirus (hMPV) is, in epidemiological terms, a relatively young addition to the catalogue of respiratory pathogens, though the burden it imposes is anything but new. First isolated from young children in the Netherlands with acute lower respiratory illness in the early 2000s, the virus is now classified as an enveloped, negative-sense, single-stranded RNA pathogen of roughly 13 kb within the Pneumovirinae subfamily of the family Pneumoviridae (Biacchesi et al., 2004). What makes hMPV worth revisiting, even two decades on, is less its taxonomy than its quiet ubiquity: surveillance from refugee settings in Kenya has placed prevalence near 5.7% (Jamal et al., 2012), while community-based estimates elsewhere range from roughly 5% to 15%, with the elderly and immunocompromised disproportionately represented among those affected (Sumino et al., 2005).
Clinically, the virus is something of a chameleon. Many infections pass as little more than a cold, a cough lingering a few days longer than expected, while others progress, sometimes unpredictably, toward bronchiolitis, pneumonia, or a syndrome resembling severe acute respiratory illness (Boivin et al., 2004). Older adults and those with compromised immunity appear to carry a disproportionate share of this severe end, a pattern documented across hospitalization-based surveillance (Falsey et al., 2003; Widmer et al., 2021), and recent case reports continue to underscore just how diagnostically elusive severe presentations can be (Filho et al., 2025). Yet no licensed vaccine or targeted antiviral currently exists; supportive care remains, more or less by default, the standard of management (Simoes et al., 2006).
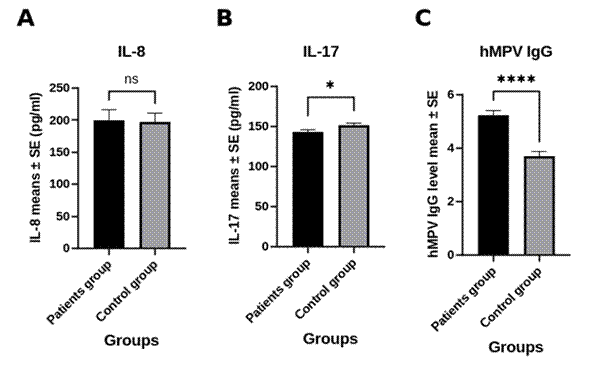
Part of the reason hMPV produces such variable disease may lie not only in the virus itself but in how forcefully, or how poorly, the host responds to it. Infection of the respiratory epithelium, mediated through the virus’s fusion (F) and attachment (G) glycoproteins, triggers pattern-recognition-receptor signaling in dendritic cells, macrophages, and epithelial cells, mobilizing a fairly stereotyped repertoire of pro-inflammatory cytokines and chemokines (Ruuskanen et al., 2011). This innate response is, in principle, protective; in practice, it can tip toward collateral damage, particularly where viral infection intersects with pre-existing airway vulnerability, as observed in asthma exacerbation (Busse et al., 2010). Among the molecules implicated in this balance, interleukin-8 (IL-8) and interleukin-17 (IL-17) have drawn particular attention.
IL-8, also known as CXCL8, behaves rather like an alarm bell for neutrophils: rapidly upregulated in infected airway epithelium and typically peaking within 12 to 24 hours of infection, it recruits neutrophils to the site of viral replication with considerable efficiency (Huck et al., 2007). The trouble is that neutrophils, once summoned, are not especially discriminating; their accumulation can clear pathogen but also injure the very tissue they are meant to defend, a dynamic implicated in both viral clearance and progressive lung injury (Alvarez et al., 2005; Medoff et al., 2009).
IL-17 tells a more ambiguous story. Secreted predominantly by Th17 cells, a lineage whose biology has been reviewed extensively elsewhere (Korn et al., 2009), it amplifies downstream inflammatory signaling and has been shown, in experimental models, to drive neutrophils into the alveolar space following hMPV-induced IL-17 release from bronchial epithelial cells via the viral G protein (Kolli et al., 2008). Yet hMPV’s cytokine signature appears genuinely distinct from that of its close relative, respiratory syncytial virus, which suggests that not every paramyxovirus infection provokes an identical Th17-skewed response, and that IL-17’s net effect, protective in some settings and pathogenic in others, may depend heavily on timing and host factors that remain incompletely mapped (Guerrero-Plata et al., 2005).
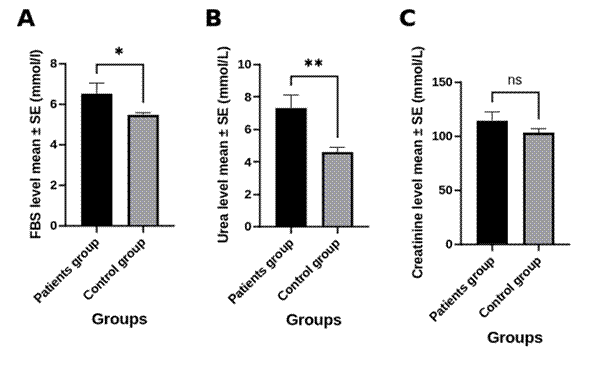
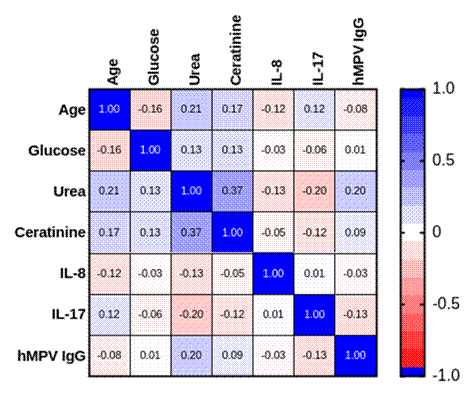
What has received comparatively little attention is whether this cytokine-driven inflammatory milieu reaches beyond the lung to influence systemic metabolism. Acute infection is well known to provoke transient hyperglycemia through counter-regulatory hormones and cytokine-mediated insulin resistance, and chronic low-grade inflammation has long been implicated in metabolic disease more broadly (Hotamisligil, 2006). Renal indices such as urea and creatinine can likewise shift with systemic inflammation or catabolic stress even absent intrinsic kidney disease. Whether hMPV infection meaningfully perturbs these parameters, and whether such perturbation might foreshadow, or merely accompany, a transient diabetogenic state, remains essentially untested, despite growing interest in how respiratory viral disease intersects with airway and systemic metabolic health (Chittiprol et al., 2025; Dickson et al., 2016). Accordingly, this case-control study set out to characterize serum IL-8, IL-17, and anti-hMPV IgG alongside fasting glucose, urea, and creatinine in hMPV-confirmed, non-diabetic patients relative to healthy controls, with the broader aim of clarifying whether this virus carries detectable implications for glycemic and renal health, including possible relevance to Type 2 diabetes risk.