3.1 Inflammatory Diseases and Bioactive Lipids in the Respiratory Tract
Cystic fibrosis (CF) lung disease has traditionally been interpreted through a largely mechanical framework, one centered on dehydrated mucus, impaired mucociliary clearance, and progressive airway obstruction. While these features unquestionably remain central to disease progression, they no longer seem sufficient to explain the extraordinary persistence of inflammation and microbial adaptation observed in many patients. Increasingly, evidence suggests that the CF airway behaves less like a passive site of obstruction and more like a highly dynamic metabolic environment where host-derived lipids, microbial metabolites, immune mediators, and environmental stress signals interact continuously. Within this altered biochemical landscape, bioactive metabolites appear to shape not only inflammatory responses but also pathogen survival, tissue remodeling, and long-term pulmonary decline (Harwood et al., 2021; Mitri et al., 2020). Figure 1 summarizes the dynamic metabolic environment of the cystic fibrosis airway, highlighting the interconnected roles of bioactive metabolites, microbial adaptation, immune dysregulation, and oxidative stress in sustaining chronic pulmonary inflammation and disease progression.
The emergence of metabolomics and breathomics has further complicated earlier assumptions about CF pathophysiology. These approaches have revealed that the respiratory tract in CF contains a remarkably diverse array of inflammatory lipids, microbial fermentation products, oxidative stress metabolites, and immunomodulatory compounds. Some of these molecules may function as biomarkers of disease activity, whereas others appear to actively drive pathology itself. The distinction, however, is not always straightforward. In many cases, metabolites simultaneously reflect and reinforce inflammation, making it difficult to separate cause from consequence. This ambiguity may partly explain why chronic airway inflammation often persists even after the introduction of highly effective CFTR modulator therapies such as elexacaftor/tezacaftor/ivacaftor (ETI) (Gramegna et al., 2026; Seidl et al., 2026).
3.2 Bioactive Lipids and Early Pulmonary Injury
Among the most intensively investigated metabolic abnormalities in CF are disruptions in sphingolipid and phospholipid metabolism. Lipids within the respiratory tract are not merely structural membrane components; they also function as signaling mediators that regulate epithelial integrity, immune cell recruitment, apoptosis, oxidative stress, and microbial defense. Under healthy conditions, these pathways remain tightly balanced. In CF, however, CFTR dysfunction appears to disturb this equilibrium at multiple levels (Lévêque et al., 2017).
One particularly important finding involves the accumulation of ceramide within airway tissues. Ceramide is a bioactive sphingolipid known to promote inflammatory signaling and epithelial injury. Experimental studies suggest that defective CFTR alters intracellular vesicular pH, impairing acid sphingomyelinase regulation and subsequently promoting ceramide accumulation (Dechecchi et al., 2011). Elevated ceramide concentrations have been associated with increased neutrophilic infiltration, oxidative stress, and heightened susceptibility to chronic bacterial colonization. Although inflammation in CF has long been linked to persistent infection, these findings imply that at least part of the inflammatory process may emerge intrinsically from disrupted lipid metabolism itself rather than solely from microbial presence (Bardin et al., 2018; Snowball et al., 2026).
The significance of these abnormalities becomes particularly evident during early disease stages. Longitudinal bronchoalveolar lavage fluid analyses in children with CF have demonstrated that specific lipid signatures can predict structural lung disease progression even before severe clinical decline becomes apparent (Horati et al., 2020). Increased ratios of long-chain ceramides to very-long-chain ceramides have been associated with worsening PRAGMA-CF scores and airway remodeling. Likewise, oxidative stress-associated lipid disturbances have been observed in young patients with relatively preserved lung function, suggesting that metabolic injury begins remarkably early in the disease process (Scholte et al., 2019).
Other lipid mediators may contribute to pulmonary injury through somewhat different mechanisms. Lysophosphatidic acid (LPA) and lysophosphatidylcholine (LPC), for example, are elevated in CF airways and appear capable of amplifying cytokine signaling, epithelial activation, and tissue remodeling responses (Horati et al., 2020). LPA signaling has been linked to amphiregulin release and neutrophil recruitment, processes that may perpetuate the chronic inflammatory cycle characteristic of CF lung disease. At the same time, protective lipid mediators such as sphingosine-1-phosphate (S1P) are often diminished. Reduced S1P levels may impair immune regulation and epithelial repair pathways, potentially worsening pulmonary damage over time (Untersmayr et al., 2020).
Interestingly, lipid dysregulation is not unique to CF and has also been reported in other inflammatory pulmonary disorders, including idiopathic pulmonary fibrosis (IPF) (Bargagli et al., 2020; Yan et al., 2017). While the pathogenic mechanisms differ substantially between these diseases, similarities in disrupted lipid signaling pathways raise the possibility that certain metabolic drivers of inflammation may transcend individual respiratory conditions. This overlap has stimulated growing interest in targeting lipid metabolism therapeutically across chronic lung diseases more broadly. Figure 2 summarizes the mechanistic relationship between CFTR dysfunction and lipid metabolic abnormalities in cystic fibrosis, highlighting how altered ceramide, LPA/LPC, and S1P signaling pathways contribute to oxidative stress, epithelial injury, neutrophilic inflammation, and progressive pulmonary decline.
3.3 Microbial Fermentation Products and Airway Inflammation
The metabolic environment of the CF airway is further complicated by microbial fermentation products generated under hypoxic and anaerobic conditions. Thick mucus plugs create oxygen gradients that favor the growth of
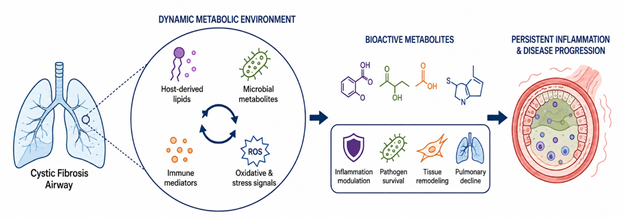
Figure 1: Metabolic and Inflammatory Interactions Driving Cystic Fibrosis Lung Disease. This figure illustrates how host-derived lipids, microbial metabolites, immune mediators, and oxidative stress signals interact within the cystic fibrosis airway microenvironment, contributing to persistent inflammation, pathogen survival, tissue remodeling, and progressive pulmonary decline.
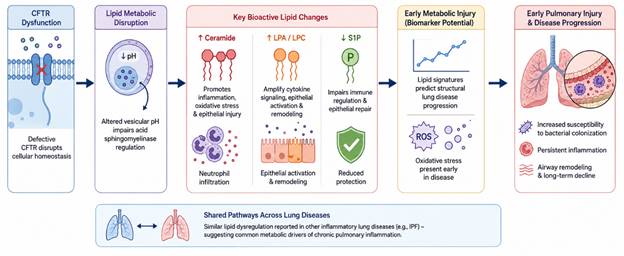
Figure 2. Bioactive Lipid Dysregulation and Early Pulmonary Injury in Cystic Fibrosis. This infographic illustrates how CFTR dysfunction disrupts lipid metabolic pathways, leading to ceramide accumulation, altered lysophospholipid signaling, oxidative stress, immune dysregulation, and progressive airway inflammation that contributes to early pulmonary injury and chronic lung disease progression in cystic fibrosis.
facultative and obligate anaerobes, many of which produce short-chain fatty acids (SCFAs) such as acetate, propionate, and butyrate. Traditionally, SCFAs have often been viewed positively because of their anti-inflammatory roles within the gastrointestinal tract. Yet within the CF lung, their effects appear considerably more complex and, at times, contradictory (Ghorbani et al., 2015).
Elevated SCFA concentrations in sputum have been associated with increased neutrophilic inflammation and heightened cytokine release, including IL-6 and IL-8 production by airway epithelial cells. Propionate, in particular, appears capable of intensifying inflammatory signaling pathways while simultaneously altering bacterial growth dynamics. What makes these metabolites especially intriguing is their concentration-dependent behavior. Lower SCFA levels may support bacterial persistence and metabolic adaptation, whereas higher acidic concentrations can inhibit certain organisms or promote the selection of resistant phenotypes (Ghorbani et al., 2015). This duality complicates simplistic interpretations of microbial metabolites as either beneficial or harmful.
The ecological implications are equally important. Anaerobic bacterial communities are increasingly recognized as active participants in CF airway disease rather than merely incidental colonizers. Shifts toward anaerobe-dominated microbial networks have been observed in patients with advanced disease or non-tuberculous mycobacterial infection (Pailhoriès et al., 2026). These organisms contribute not only to local metabolite production but also to broader alterations in airway pH, redox balance, and inflammatory signaling. In effect, the microbial community continuously reshapes the biochemical conditions that sustain its own persistence.
3.4 Immunometabolism and Pathogen Adaptation
Perhaps one of the most fascinating developments in recent years involves the concept of immunometabolism in CF. Activated immune cells do not simply produce inflammatory cytokines; they also undergo profound metabolic reprogramming that can influence pathogen behavior directly. Neutrophils and macrophages in CF airways exhibit altered glycolytic activity and increased production of metabolites such as succinate and itaconate (Makam et al., 2009).
Pseudomonas aeruginosa, the dominant chronic pathogen in many adults with CF, appears exceptionally well adapted to exploit these metabolic conditions. Rather than relying solely on traditional nutrients, P. aeruginosa preferentially consumes host-derived succinate as a carbon source, facilitating rapid growth and promoting biofilm-associated phenotypes (Palmer & Whiteley, 2015; Riquelme & Prince, 2021). The organism’s metabolic flexibility likely contributes substantially to its extraordinary persistence within CF airways.
Even more striking is the relationship between P. aeruginosa and itaconate. Normally, itaconate functions as an anti-inflammatory immunometabolite intended to suppress excessive inflammation and inhibit bacterial metabolism. However, P. aeruginosa possesses metabolic pathways capable of degrading and utilizing itaconate, effectively converting a host defense mechanism into a survival advantage (Riquelme & Prince, 2021). This phenomenon highlights the sophistication of metabolic host-pathogen interactions in chronic CF infection. The bacteria are not merely surviving within inflammation; they are adapting specifically to the inflammatory metabolites generated by the host immune response itself.
3.5 The Gut–Lung Axis and Systemic Metabolic Signaling
Although CF lung disease remains the major determinant of morbidity and mortality, growing evidence suggests that pulmonary inflammation cannot be fully understood without considering systemic microbial interactions, particularly those involving the gut microbiome. CF-associated intestinal dysbiosis arises from multiple factors, including altered epithelial ion transport, frequent antibiotic exposure, pancreatic insufficiency, and specialized nutritional regimens (Thavamani et al., 2021).
This dysbiosis may influence respiratory disease through the gut–lung axis, a bidirectional communication network mediated partly by microbial metabolites. Indole derivatives produced during bacterial tryptophan metabolism have attracted particular interest because they serve as ligands for the aryl hydrocarbon receptor (AHR), an important regulator of epithelial integrity and immune homeostasis (Hou et al., 2025). Elevated indole-related compounds in exhaled breath have been associated with poorer pulmonary outcomes and chronic P. aeruginosa colonization.
The implications of these findings remain somewhat
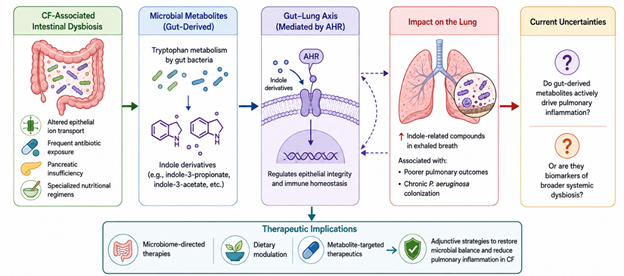
Figure 3. Gut–Lung Axis and Microbial Metabolite Signaling in Cystic Fibrosis. This Figure illustrates how CF-associated intestinal dysbiosis may influence pulmonary inflammation through gut-derived microbial metabolites, particularly indole derivatives that regulate epithelial integrity and immune homeostasis via aryl hydrocarbon receptor (AHR)-mediated signaling pathways.
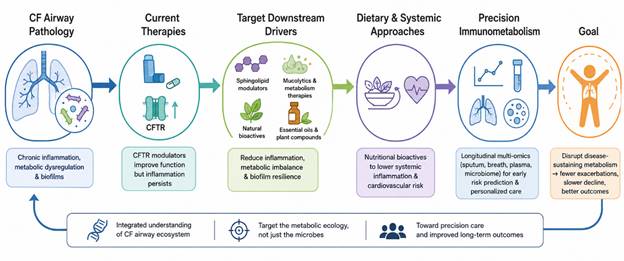
Figure 4. Precision Immunometabolism-Based Therapeutic Framework for Cystic Fibrosis Lung Disease. This figure illustrates how persistent airway inflammation and metabolic dysregulation continue despite CFTR modulator therapy, highlighting emerging strategies targeting sphingolipid signaling, biofilm metabolism, natural bioactives, and precision metabolomic profiling to improve long-term clinical outcomes in cystic fibrosis.
uncertain. It is still unclear whether gut-derived metabolites actively contribute to pulmonary inflammation or primarily function as biomarkers reflecting broader systemic dysbiosis. Nevertheless, mounting evidence suggests that intestinal microbial disturbances may influence pulmonary immunity more substantially than previously appreciated (Marsh et al., 2026). This has prompted increasing interest in microbiome-directed interventions, dietary modulation, and metabolite-targeted therapeutics as adjunctive strategies in CF care. Figure 3 summarizes the proposed gut–lung axis in cystic fibrosis, highlighting the role of intestinal dysbiosis, tryptophan-derived microbial metabolites, and AHR signaling in modulating pulmonary immunity, chronic inflammation, and susceptibility to persistent respiratory colonization.
3.6 Emerging Therapeutic Strategies Beyond CFTR Correction
The development of CFTR modulators has undoubtedly transformed CF management. ETI therapy improves lung function, nutritional status, and survival in many patients. Yet despite these remarkable advances, complete resolution of airway inflammation and chronic infection remains uncommon (Harwood et al., 2021). In many individuals, inflammatory pathways appear to persist despite partial restoration of CFTR activity, suggesting that chronic metabolic alterations may become somewhat self-sustaining over time.
Consequently, attention has shifted toward therapies targeting downstream inflammatory and metabolic drivers. Sphingolipid modulators, including acid sphingomyelinase inhibitors such as amitriptyline, have shown potential for reducing ceramide accumulation and inflammatory signaling (Dechecchi et al., 2011; Mitri et al., 2020). Likewise, mucolytic and metabolism-based therapies aimed at disrupting biofilm nutrient networks continue to evolve. Natural bioactive compounds have also emerged as promising adjunctive candidates. Curcumin, resveratrol, and baicalin possess anti-inflammatory and antioxidant properties that may influence CF-related signaling pathways (Chang et al., 2021; Luo et al., 2025). Baicalin, for example, has demonstrated anti-fibrotic and metabolic regulatory effects in pulmonary injury models through modulation of glutathione and lipid metabolism. Essential oils and plant-derived bioactive substances have similarly shown anti-biofilm activity against Staphylococcus aureus isolates obtained from CF patients (Papa et al., 2020). Although many of these approaches remain experimental, they reflect a broader conceptual shift toward targeting the metabolic ecology of CF rather than focusing exclusively on antimicrobial eradication.
At the same time, interest is growing in dietary and nutritional bioactives capable of reducing systemic inflammation and cardiovascular complications associated with CF (Trandafir et al., 2023). These interventions may become increasingly relevant as life expectancy improves and long-term comorbidities emerge more prominently.
The understanding of CF lung disease has evolved considerably from earlier models centered solely on mucus obstruction and bacterial colonization. Current evidence increasingly portrays the CF airway as a metabolically active ecosystem shaped by intricate interactions among bioactive lipids, microbial fermentation products, immune signaling metabolites, oxidative stress pathways, and systemic microbial communication networks. Yet many uncertainties remain unresolved. It is still difficult to determine which metabolites primarily drive pathology and which merely reflect ongoing disease activity. Figure 4 summarizes the evolving therapeutic paradigm of cystic fibrosis (CF) lung disease, emphasizing the transition from conventional CFTR correction toward precision immunometabolism approaches that target chronic inflammation, metabolic imbalance, microbial persistence, and systemic complications associated with CF progression.
Nevertheless, the concept of “precision immunometabolism” is beginning to emerge as a potentially transformative framework for future CF management. Longitudinal metabolomic profiling of sputum, breath condensates, plasma, and microbial communities may eventually help identify patients at risk for exacerbation, treatment failure, or accelerated structural decline before irreversible injury occurs. Importantly, the future of CF therapy may depend not only on correcting CFTR dysfunction but also on disrupting the metabolic conditions that allow chronic inflammation and microbial persistence to endure.
