1. Introduction
Microorganisms play a fundamental role in shaping human physiology, immune development, and disease susceptibility. Rather than existing as isolated organisms, microbes form highly structured communities—collectively known as microbiomes—that inhabit different anatomical niches including the oral cavity, gastrointestinal tract, skin, and respiratory system (Wade, 2013; Kilian et al., 2016). These microbial ecosystems are dynamic and respond continuously to environmental conditions, host immunity, and microbial interactions. Under balanced conditions, microbial communities contribute to physiological homeostasis and host protection. However, disturbances in microbial composition or function can lead to dysbiosis, a state of ecological imbalance that has been increasingly linked to chronic inflammatory and systemic diseases.
Among the various microbial ecosystems of the human body, the oral microbiome represents one of the most diverse and densely populated microbial communities. More than 700 bacterial species have been identified in the oral cavity, along with fungi, viruses, and archaea that colonize mucosal surfaces, dental plaques, and gingival tissues (Dewhirst et al., 2010; Deo & Deshmukh, 2019). These microorganisms exist in complex multispecies biofilms where metabolic cooperation, competition, and signaling determine community structure and stability. In healthy individuals, the oral microbiome remains in a dynamic equilibrium maintained through host immune responses, saliva composition, and interspecies microbial interactions (Kilian et al., 2016). When this equilibrium is disrupted, however, pathogenic microbial consortia may emerge and initiate inflammatory disease processes.
Oral dysbiosis represents a functional shift in microbial communities rather than simply the presence of pathogenic species. This shift involves altered metabolic activity, increased virulence factor expression, and dysregulated host–microbe interactions (Hajishengallis, 2014). Periodontal diseases exemplify dysbiosis-driven conditions, where microbial biofilms trigger chronic inflammation and progressive destruction of tooth-supporting tissues (Lasserre et al., 2018). Biofilms provide structural protection to resident microbes by embedding them within extracellular polymeric matrices that reduce susceptibility to antimicrobial agents and host immune responses (Costerton et al., 1999; Muhammad et al., 2020). As a result, infections associated with biofilms are often persistent and difficult to eradicate using conventional antimicrobial therapies.
The complexity of polymicrobial biofilms further contributes to disease progression through mechanisms of polymicrobial synergy and immune modulation. Certain keystone pathogens, such as Porphyromonas gingivalis, can alter the microbial community composition and manipulate host immune responses despite being present in relatively low abundance (Hajishengallis & Lamont, 2012). This phenomenon highlights the importance of community-level microbial interactions rather than the pathogenicity of individual microorganisms alone. Persistent activation of inflammatory pathways subsequently contributes to tissue damage and systemic immune dysregulation (Cekici et al., 2014).
Beyond localized oral diseases, dysbiotic microbial communities have increasingly been implicated in systemic pathologies. Microbial dissemination from the oral cavity can occur through hematogenous or enteral routes, allowing oral microorganisms or their inflammatory components to reach distant organs (Kitamoto et al., 2020; Khor et al., 2021). Routine activities such as chewing or tooth brushing may introduce oral bacteria into the bloodstream, particularly in individuals with periodontal inflammation (Parahitiyawa et al., 2009). Once circulating, bacterial components including lipopolysaccharides activate innate immune receptors such as Toll-like receptors, triggering systemic inflammatory cascades (Jang et al., 2015; Keller et al., 2011).
Increasing evidence suggests that oral dysbiosis contributes to the development of several systemic diseases, including cardiovascular, autoimmune, and neurodegenerative disorders. Chronic periodontal inflammation has been associated with endothelial dysfunction and atherosclerotic plaque formation, thereby elevating cardiovascular risk (Aarabi et al., 2018). Similarly, autoimmune conditions such as rheumatoid arthritis have been linked to periodontal pathogens capable of inducing protein citrullination and immune tolerance breakdown (Konig et al., 2016). In neurodegenerative diseases including Parkinson’s disease, bacterial inflammagens and inflammatory mediators derived from dysbiotic microbial communities have been detected in systemic circulation and brain tissues, suggesting a potential microbial contribution to neuroinflammatory processes (Adams et al., 2019).
The persistence of microbial biofilms and the growing prevalence of antimicrobial resistance highlight the need for innovative therapeutic strategies capable of disrupting pathogenic microbial communities while minimizing systemic toxicity. Conventional antibiotics often demonstrate limited efficacy against biofilm-associated infections and may contribute to the emergence of resistant strains. Consequently, research has increasingly focused on alternative antimicrobial approaches, including antimicrobial peptides and nanomaterial-based therapeutics, which offer promising mechanisms for combating microbial pathogens.
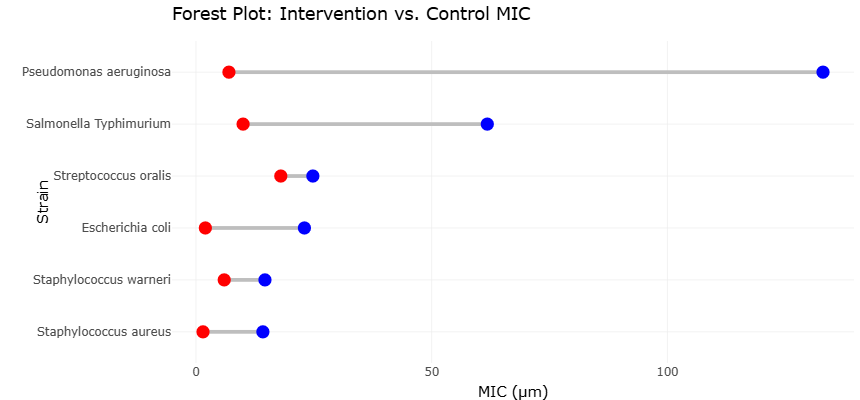
Antimicrobial peptides represent a class of naturally occurring defense molecules capable of targeting bacterial membranes and disrupting microbial viability. Recent investigations have demonstrated the potent antibacterial activity of synthetic or naturally derived peptides against both Gram-negative and Gram-positive pathogens. Comparative analyses of minimum inhibitory concentration (MIC) values illustrate the effectiveness of these compounds relative to conventional antibiotics. For example, the antimicrobial peptide SQQ30 has demonstrated strong inhibitory activity against pathogens such as Escherichia coli, Pseudomonas aeruginosa, and Staphylococcus aureus, often at substantially lower concentrations than traditional antibiotics like gentamicin (Di Napoli et al., 2024). These comparative antimicrobial profiles, the potential of peptide-based therapeutics as effective tools for controlling pathogenic microbial populations associated with dysbiosis and infection.
In parallel with peptide-based antimicrobials, nanotechnology has emerged as a powerful platform for developing next-generation antimicrobial agents. Metallic nanoparticles, particularly silver nanoparticles (AgNPs), exhibit broad-spectrum antimicrobial properties resulting from their ability to disrupt bacterial membranes, generate reactive oxygen species, and interfere with cellular metabolic processes. Recent studies have explored environmentally friendly synthesis methods for AgNPs using plant extracts, producing nanoparticles with enhanced biological compatibility and antimicrobial performance (Syafiuddin et al., 2018). These green-synthesized nanoparticles have demonstrated measurable antibacterial activity against environmental and clinically relevant bacterial strains.
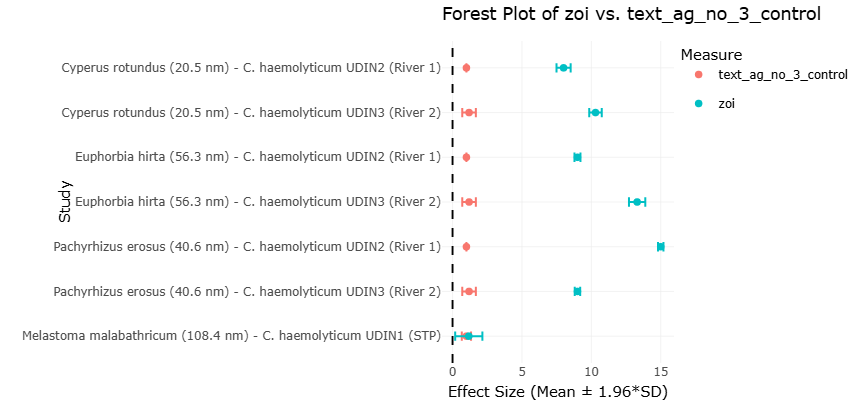
Plant-mediated silver nanoparticles exhibit varying degrees of antibacterial efficacy depending on synthesis source, particle size, and microbial strain. For example, nanoparticles synthesized using plant extracts such as Cyperus rotundus, Euphorbia hirta, and Pachyrhizus erosus have shown measurable zones of inhibition against Chromobacterium haemolyticum isolates, significantly exceeding the inhibitory effects observed with silver nitrate controls (Syafiuddin et al., 2018). These results suggest that nanoparticle-based antimicrobial systems may offer effective strategies for controlling microbial populations in both environmental and clinical contexts.
The integration of antimicrobial peptides and nanomaterials represents a promising frontier in microbial control strategies. Nanoparticles can enhance antimicrobial delivery, increase stability of bioactive compounds, and facilitate targeted interactions with microbial cells. Moreover, the physicochemical properties of nanoparticles—including particle size, surface charge, and functionalization—can be optimized to improve antimicrobial activity against resistant microbial communities embedded within biofilms. Such properties are particularly relevant in the context of oral microbial dysbiosis, where biofilm resilience often limits the effectiveness of conventional therapies.
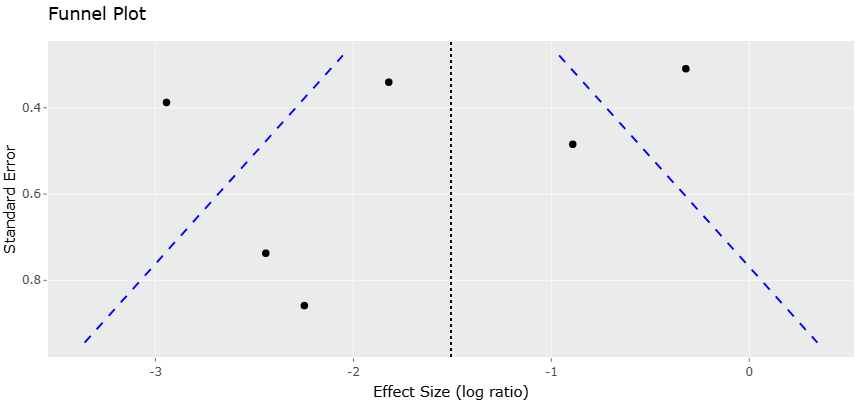
Despite growing interest in nanoparticle-based antimicrobials, considerable variability exists across studies regarding nanoparticle synthesis methods, antimicrobial assays, and microbial targets. Differences in particle size, plant-derived reducing agents, and experimental conditions can influence antimicrobial performance, complicating direct comparisons across studies. Similarly, studies evaluating antimicrobial peptides and nanoparticle formulations often employ different microbial strains and assay methodologies, resulting in heterogeneous findings.
Given these variations, systematic evidence synthesis is essential to better understand the antimicrobial potential of emerging biomaterials and their implications for microbial dysbiosis and infection control. Integrating data from multiple studies enables identification of consistent antimicrobial patterns, comparative efficacy across antimicrobial agents, and potential mechanisms underlying microbial susceptibility. Such analyses are particularly important for evaluating innovative antimicrobial strategies designed to overcome biofilm resistance and microbial community complexity.
Therefore, this systematic review aims to synthesize existing evidence on microbial dysbiosis, antimicrobial interventions, and nanoparticle-based antimicrobial strategies. By integrating findings from microbiome studies, antimicrobial peptide research, and nanoparticle-based antibacterial investigations, this study seeks to clarify how emerging antimicrobial technologies may contribute to controlling dysbiotic microbial communities and reducing systemic disease risk. Understanding the interplay between microbial ecology, host immunity, and advanced antimicrobial technologies is critical for developing effective therapeutic strategies capable of restoring microbial balance and improving human health outcomes.