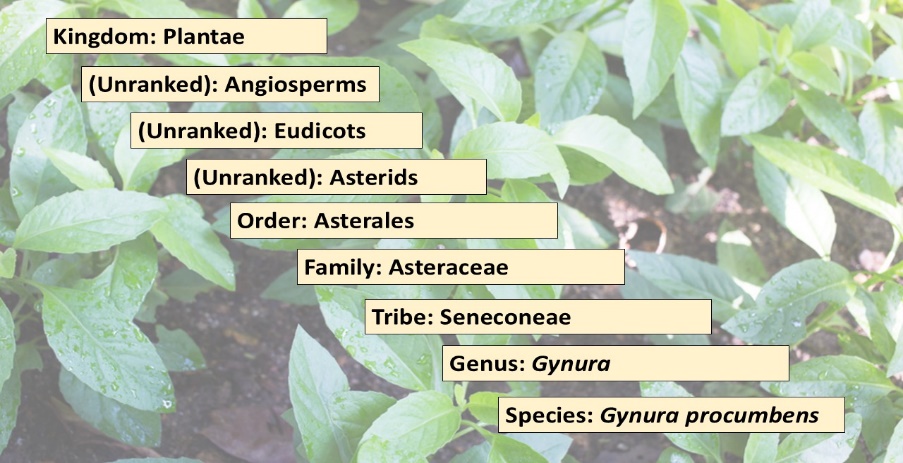
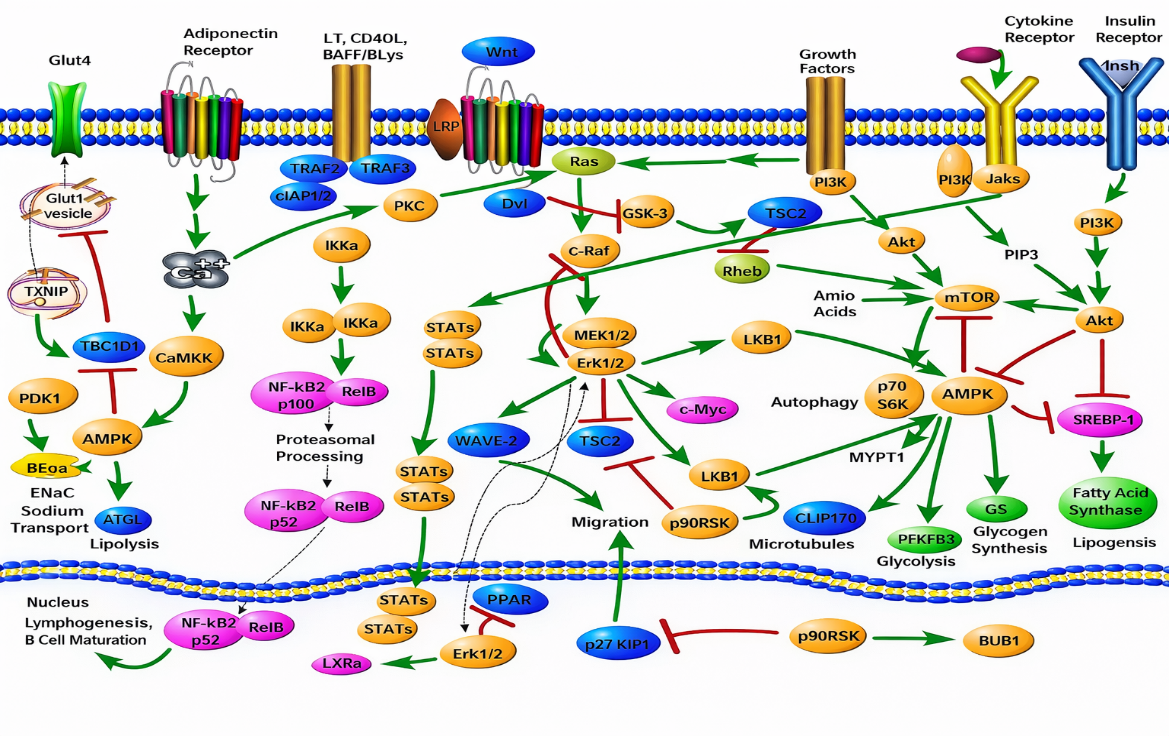
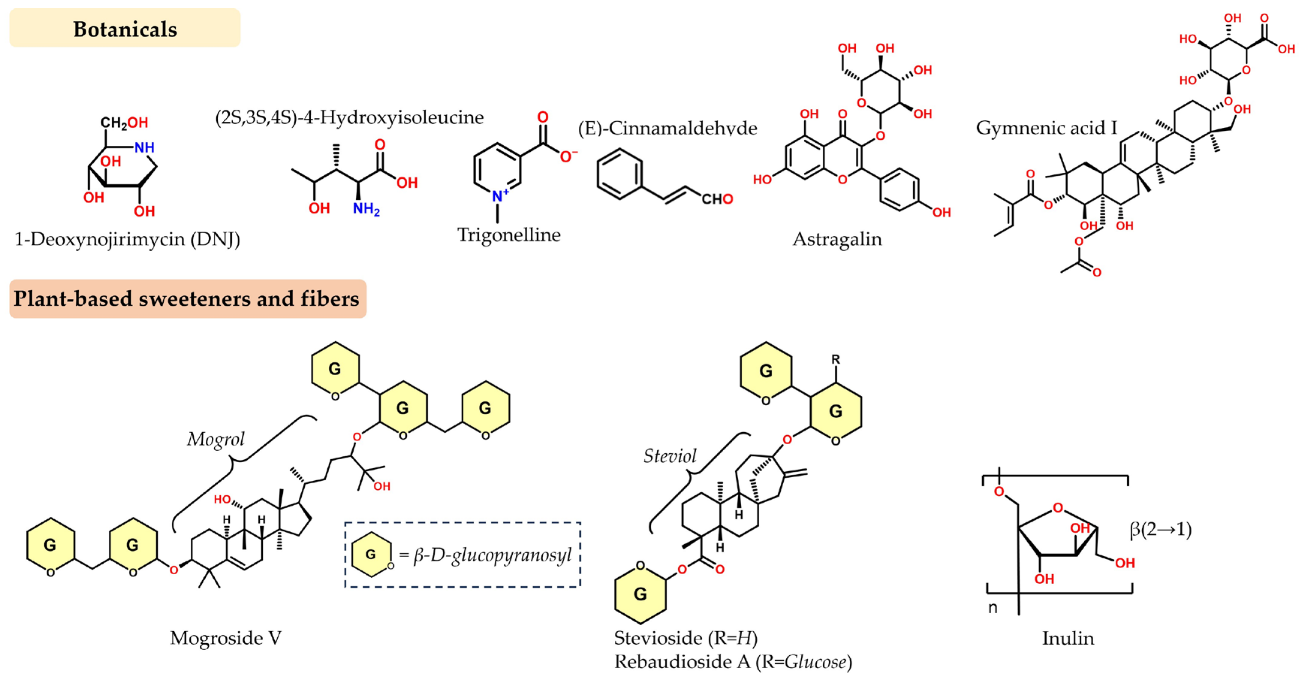
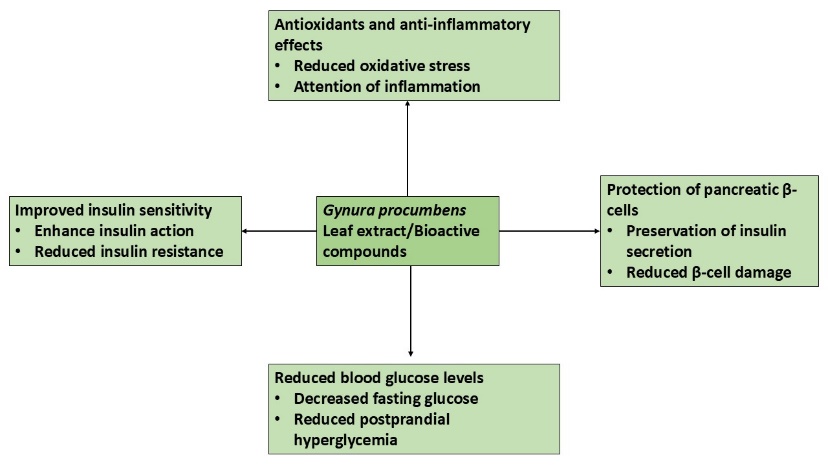
Akmar, K., & Noor, M. M. (2020). The potential effect of Gynura procumbens aqueous extract as anti-hyperglycaemia, pro-fertility and libido agent towards diabetes-induced male rats. In Proceeding International Conference on Science and Engineering.
Alfheeaid, H. A., et al. (2023). Effect of various intermittent fasting protocols on hyperglycemia-induced cognitive dysfunction in rats. Brain Sciences, 13(2), 165.
Algariri, K., Atangwho, I. J., Asmawi, M. Z., Sadikun, A., Murugaiyah, V., & Ismail, N. (2013). Hypoglycemic and anti-hyperglycemic study of Gynura procumbens leaf extracts. Asian Pacific Journal of Tropical Biomedicine, 3(5), 358–366.
Algariri, K., Meng, K. Y., Atangwho, I. J., Asmawi, M. Z., Sadikun, A., Murugaiyah, V., & Ismail, N. (2014). Antihyperglycaemic and toxicological evaluations of extract and fractions of Gynura procumbens leaves. Tropical Life Sciences Research, 25(1), 75–93.
Almatroudi, A., et al. (2023). Effects and mechanisms of kaempferol in the management of cancers through modulation of inflammation and signal transduction pathways. International Journal of Molecular Sciences, 24(10), 8630.
Amin, M. Z., et al. (2021). Assessment of antibacterial, anti-inflammatory, and cytotoxic effects of different extracts of Gynura procumbens leaf. Current Therapeutic Research, 95, 100636.
Anjum, J., et al. (2024). Exploration of the GLUT-4 modulation potential of Gynura procumbens as a mechanism behind its antidiabetic activity. Biomedical and Pharmacology Journal, 17(4), 2769–2778.
Aung, C. L., et al. (2021). Blood glucose-lowering effect of water and ethanolic extracts of Gynura procumbens (Lour.) Merr. Traditional & Kampo Medicine, 8(2), 138–147.
Badsha, S. A., et al. (2024). Effect of Gynura procumbens leaf extract with biological nanoparticles on streptozotocin-induced hyperglycemia in a rat model. Journal of Advanced Biotechnology and Experimental Therapeutics, 7(1), 255–265.
Bailey, C. J., & Day, C. (2004). Metformin: Its botanical background. Practical Diabetes International, 21(3), 115–117.
Bakar, M. A., Deb, N., & Rahman, M. A. (2018). Advancing medical science through nanobiotechnology: Prospects, applications, and future directions. Journal of Primeasia, 1(1), 1–10.
Balwan, W. K., Saba, N., & Zargar, J. I. (2022). Burden of diabetes and role of medicinal plants in its treatment. Saudi Journal of Medical and Pharmaceutical Sciences, 8(7), 355–361.
Blonde, L., San Juan, Z. T., & Bolton, P. (2014). Fixed-dose combination therapy in type 2 diabetes mellitus. Endocrine Practice, 20(12), 1322–1332.
Bodeker, G., & Shekar, S. (2009). Health and beauty from the rainforest: Malaysian traditions of ramuan. Biotropics Ramuan.
Borsha, R. P., et al. (2020). Study of steroid induced hyperglycemia against traditional hypoglycemic plant extracts in rat model. Asian Journal of Pharmacy and Pharmacology, 6(4), 254–260.
Cao, M.-M., et al. (2025). Astragalin alleviates lipopolysaccharide-induced depressive-like behavior in mice by preserving blood-brain barrier integrity and suppressing neuroinflammation. Free Radical Biology and Medicine, 232, 340–352.
Choi, S.-I., Park, M. H., & Han, J.-S. (2016). Gynura procumbens extract alleviates postprandial hyperglycemia in diabetic mice. Preventive Nutrition and Food Science, 21(3), 181–186.
Chu, T., et al. (2024). Kaempferol regulating macrophage foaming and atherosclerosis through Piezo1-mediated MAPK/NF-κB and Nrf2/HO-1 signaling pathway. Journal of Advanced Research.
Dong, R., et al. (2023). Rutin alleviates EndMT by restoring autophagy through inhibiting HDAC1 via PI3K/AKT/mTOR pathway in diabetic kidney disease. Phytomedicine, 112, 154700.
Gansau, J. A., et al. (2012). Hypoglycemic effects of Gynura procumbens fractions on streptozotocin-induced diabetic rats involved phosphorylation of GSK3β (Ser-9) in liver. Sains Malaysiana, 41, 969–975.
Gershenzon, J., & Dudareva, N. (2007). The function of terpene natural products in the natural world. Nature Chemical Biology, 3(7), 408–414.
Guo, H., et al. (2021). Quercetin induces pro-apoptotic autophagy via SIRT1/AMPK signaling pathway in human lung cancer cell lines A549 and H1299 in vitro. Thoracic Cancer, 12(9), 1415–1422.
Guo, S., et al. (2021). Exploring the protective effect of Gynura procumbens against type 2 diabetes mellitus by network pharmacology and validation in C57BL/KsJ db/db mice. Food & Function, 12(4), 1732–1744.
Han, S.-H., et al. (2022). Myricetin induces apoptosis and autophagy in human gastric cancer cells through inhibition of the PI3K/Akt/mTOR pathway. Heliyon, 8(5).
Hassan, Z., Yam, M. F., Ahmad, M., & Yusof, A. P. M. (2010). Antidiabetic properties and mechanism of action of Gynura procumbens water extract in streptozotocin-induced diabetic rats. Molecules, 15(12), 9008–9023.
Holman, N., Young, B., & Gadsby, R. (2015). Invited editorial: Current prevalence of Type 1 and Type 2 diabetes in adults and children in the UK. Diabetic Medicine, 32, 1119–1120.
Hou, D.-D., et al. (2022). Therapeutic effects of myricetin on atopic dermatitis in vivo and in vitro. Phytomedicine, 102, 154200.
Hu, N., et al. (2022). Comparative evaluation of the effect of metformin and insulin on gut microbiota and metabolome profiles of type 2 diabetic rats induced by the combination of streptozotocin and high-fat diet. Frontiers in Pharmacology, 12, 794103.
Huang, D., et al. (2024). Myricetin ameliorates airway inflammation and remodeling in asthma by activating Sirt1 to regulate the JNK/Smad3 pathway. Phytomedicine, 135, 156044.
Huang, Q., et al. (2025). Global burden and risk factors of type 2 diabetes mellitus from 1990 to 2021, and forecasts to 2050. Frontiers in Endocrinology, 16, 1538143.
Jacob, B., & Narendhirakannan, R. (2019). Role of medicinal plants in the management of diabetes mellitus: A review. 3 Biotech, 9(1), 4.
Jang, H.-Y., et al. (2023). Fisetin inhibits UVA-induced expression of MMP-1 and MMP-3 through the NOX/ROS/MAPK pathway in human dermal fibroblasts and human epidermal keratinocytes. International Journal of Molecular Sciences, 24(24), 17358.
Jobaer, M. A., et al. (2023). Phytochemical and biological investigation of an indigenous plant of Bangladesh, Gynura procumbens (Lour.) Merr.: Drug discovery from nature. Molecules, 28(10), 4186.
Kaewseejan, N., Puangpronpitag, D., & Nakornriab, M. (2012). Evaluation of phytochemical composition and antibacterial property of Gynura procumbens extract.
Kaewseejan, N., Sutthikhum, V., & Siriamornpun, S. (2015). Potential of Gynura procumbens leaves as source of flavonoid-enriched fractions with enhanced antioxidant capacity. Journal of Functional Foods, 12, 120–128.
Ketuly, K. (2011). Acute toxicity study and wound healing potential of Gynura procumbens leaf extract in rats. Journal of Medicinal Plants Research, 5(12), 2551–2558.
Khatun, A., et al. (2024). Tanshinone IIA as a promising natural inhibitor of the STING pathway: A computational exploration toward neuroinflammatory therapy. Australian Herbal Insight, 7(1), 1–13.
Kim, H. H., et al. (2021). Potential antioxidant and anti-inflammatory function of Gynura procumbens polyphenols ligand. International Journal of Molecular Sciences, 22(16), 8716.
Ko, C.-H., et al. (2005). Myricetin inhibits matrix metalloproteinase 2 protein expression and enzyme activity in colorectal carcinoma cells. Molecular Cancer Therapeutics, 4(2), 281–290.
Kooti, W., et al. (2016). The role of medicinal plants in the treatment of diabetes: A systematic review. Electronic Physician, 8(1), 1832.
Krishnan, V., Ahmad, S., & Mahmood, M. (2015). Antioxidant potential in different parts and callus of Gynura procumbens and different parts of Gynura bicolor. BioMed Research International, 2015, 147909.
Lee, H.-W., et al. (2012). Antidiabetic effect of Gynura procumbens leaves extracts involve modulation of hepatic carbohydrate metabolism in streptozotocin-induced diabetic rats. Journal of Medicinal Plants Research, 6(5), 796–812.
Levetan, C. (2007). Oral antidiabetic agents in type 2 diabetes. Current Medical Research and Opinion, 23(4), 945–952.
Li, H., et al. (2023). Kaempferol prevents acetaminophen-induced liver injury by suppressing hepatocyte ferroptosis via Nrf2 pathway activation. Food & Function, 14(4), 1884–1896.
Li, N., et al. (2023). Kaempferol attenuates gouty arthritis by regulating the balance of Th17/Treg cells and secretion of IL-17. Inflammation, 46(5), 1901–1916.
Li, Q., et al. (2025). Astragalin exerted hypoglycemic effect by both inhibiting α-glucosidase and modulating AMPK signaling pathway. Nutrients, 17(3), 406.
Li, W., et al. (2017). Astragalin reduces hexokinase 2 through increasing miR-125b to inhibit the proliferation of hepatocellular carcinoma cells in vitro and in vivo. Journal of Agricultural and Food Chemistry, 65(29), 5961–5972.
Liu, Y., et al. (2024). Rutin ameliorated lipid metabolism dysfunction of diabetic NAFLD via AMPK/SREBP1 pathway. Phytomedicine, 126, 155437.
Lu, X.-L., et al. (2017). Quercetin attenuates high fructose feeding-induced atherosclerosis by suppressing inflammation and apoptosis via ROS-regulated PI3K/AKT signaling pathway. Biomedicine & Pharmacotherapy, 85, 658–671.
Luo, X., et al. (2022). The protective effect of quercetin on macrophage pyroptosis via TLR2/Myd88/NF-κB and ROS/AMPK pathway. Life Sciences, 291, 120064.
Mahmood, A., et al. (2010). Anti-ulcerogenic activity of Gynura procumbens leaf extract against experimentally-induced gastric lesions in rats. Journal of Medicinal Plants Research, 4(8), 685–691.
Meng, X., et al. (2019). A kaempferol-3-O-β-d-glucoside, intervention effect of astragalin on estradiol metabolism. Steroids, 149, 108413.
Mohiuddin, G. S., et al. (2019). Uncommon side effects of commonly used anti-diabetics: Time to monitor them. International Journal of Pharmaceutical Sciences and Research, 10, 4145–4148.
Mou, K. M., & Dash, P. R. (2016). A comprehensive review on Gynura procumbens leaves. International Journal of Pharmacognosy, 3(4), 167–174.
Nasiruddin, M., & Sinha, S. N. (2020). Phytochemical screening and antioxidant, antibacterial efficacy of Gynura procumbens (Lour.) Merr. Asian Journal of Medical and Biological Research, 6(2), 187–195.
Pai, P. Y., et al. (2019). Anti-diabetic effects of Gynura bicolor aqueous extract in mice. Journal of Food Science, 84(6), 1631–1637.
Pandey, P., et al. (2021). Rutin (bioflavonoid) as cell signaling pathway modulator: Prospects in treatment and chemoprevention. Pharmaceuticals, 14(11), 1069.
Perry, L. M., & Metzger, J. (1980). Medicinal plants of East and Southeast Asia: Attributed properties and uses.
Rahman, A. M., & Asad, M. A. (2013). Chemical and biological investigations of the leaves of Gynura procumbens.
Robiul, M., et al. (2023). In silico drug-likeness and safety profiling of tinosporaside: A natural alternative to celecoxib for COX-2 inhibition. Journal of Primeasia, 4(1), 1–11.
Rui, L. (2014). Energy metabolism in the liver. Comprehensive Physiology, 4(1), 177–197.
Sampath, C., et al. (2020). Inhibition of GSK-3β restores delayed gastric emptying in obesity-induced diabetic female mice. American Journal of Physiology-Gastrointestinal and Liver Physiology, 319(4), G481–G493.
Sathiyaseelan, A., et al. (2021). Evaluation of phytochemicals, antioxidants, and antidiabetic efficacy of various solvent fractions of Gynura procumbens (Lour.) Merr. Process Biochemistry, 111, 51–62.
Shohel, M. A., et al. (2024). Anthelmintic potential of discarded Litchi chinensis seeds: A sustainable approach to agricultural by-product utilization. Journal of Pharmacopuncture, 27(4), 340.
Shohel, M. A., et al. (2024). Natural diabetes treatment with Litchi seeds extract in vivo. Journal of Angiotherapy, 8(7), 1–13.
Siriamornpun, S., et al. (2021). Characterization of polysaccharides from Gynura procumbens with relation to their antioxidant and anti-glycation potentials. Biocatalysis and Agricultural Biotechnology, 32, 101957.
Situmorang, P. C., et al. (2025). Effect of Gynura procumbens (Lour.) Merr. on the histopathology of the diabetic pancreas via caspase family expression. Journal of Pharmacy and Pharmacognosy Research, 13(3), 836–847.
Srivastava, A., et al. (2023). Potential of quercetin to protect cadmium induced cognitive deficits in rats by modulating NMDA-R mediated downstream signaling and PI3K/AKT–Nrf2/ARE signaling pathways in hippocampus. NeuroMolecular Medicine, 25(3), 426–440.
Sun, G. Y., et al. (2015). Quercetin attenuates inflammatory responses in BV-2 microglial cells: Role of MAPKs on the Nrf2 pathway and induction of heme oxygenase-1. PLOS ONE, 10(10), e0141509.
Sun, M.-Y., et al. (2023). Astragalin ameliorates renal injury in diabetic mice by modulating mitochondrial quality control via AMPK-dependent PGC1α pathway. Acta Pharmacologica Sinica, 44(8), 1676–1686.
Sun, X.-Y., et al. (2021). Rutin prevents tau pathology and neuroinflammation in a mouse model of Alzheimer’s disease. Journal of Neuroinflammation, 18(1), 131.
Tahsin, M. R., et al. (2022). In vivo and in silico assessment of diabetes ameliorating potentiality and safety profile of Gynura procumbens leaves. Evidence-Based Complementary and Alternative Medicine, 2022, 9095504.
Tan, H.-L., et al. (2016). Gynura procumbens: An overview of the biological activities. Frontiers in Pharmacology, 7, 52.
Tan, J. N., et al. (2020). Antioxidant and anti-inflammatory effects of genus Gynura: A systematic review. Frontiers in Pharmacology, 11, 504624.
Tang, X.-L., et al. (2015). Protective effect of kaempferol on LPS plus ATP-induced inflammatory response in cardiac fibroblasts. Inflammation, 38(1), 94–101.
Uthia, R., et al. (n.d.). Activity of ethanol extract of Gynura procumbens (Lour.) Merr. leaf to decrease blood glucose level and recover pancreatic histopathology in white male mice induced by alloxan.
Wang, Z., et al. (2021). The flavonoid astragalin shows anti-tumor activity and inhibits PI3K/AKT signaling in gastric cancer. Chemical Biology & Drug Design, 98(5), 779–786.
Wentling, G., & Kim, E. (2017). Glucophage® (metformin hydrochloride), the wonder drug: A biguanide class treatment of type 2 diabetes. Monarch Review, 4, 112–128.
Wong, S. K., et al. (2015). Anti-malarial and anti-inflammatory effects of Gynura procumbens are mediated by kaempferol via inhibition of glycogen synthase kinase-3β (GSK3β). Sains Malaysiana, 44(10), 1489–1500.
Xu, J., et al. (2024). Quercetin inhibited LPS-induced cytokine storm by interacting with the AKT1-FoxO1 and Keap1-Nrf2 signaling pathway in macrophages. Scientific Reports, 14(1), 20913.
Yam, M. F., et al. (2009). Toxicology evaluation of standardized methanol extract of Gynura procumbens. Journal of Ethnopharmacology, 123(2), 244–249.
Yu, J., et al. (2024). Genetic differences among populations of Gynura procumbens (Lour.) Merr. Genetic Resources and Crop Evolution, 71(5), 1687–1693.
Zeb, A., et al. (2026). Exploring the anticancer potential of astragalin in triple negative breast cancer cells by attenuating glycolytic pathway through AMPK/mTOR. Current Medicinal Chemistry, 33(7), 1405–1418.
Zhao, X., Chen, X., & Yue, C. (2024). Rutin ameliorates inflammation and oxidative stress in ulcerative colitis by inhibiting NLRP3 inflammasome signaling pathway. Cell Biochemistry and Biophysics, 82(4), 3715–3726.
Zhu, M.-L., et al. (2020). Myricetin induces apoptosis and autophagy by inhibiting PI3K/Akt/mTOR signalling in human colon cancer cells. BMC Complementary Medicine and Therapies, 20(1), 209.
Ziaul Amin, M., et al. (2021). Evaluation of medicinal effects of Gynura procumbens leave extracts on oxidative, glycemic, lipidomics, and enzymatic profiles in alloxan-induced diabetic mice. Journal of Diabetes & Metabolism, 12, 876.